

A Role for Plasmacytoid Dendritic Cells in Lupus Save

Know-it-now
- Plasmacytoid dendritic cells (pDCs) are central to lupus pathogenesis via TLR7/9- and cGAS/STING-driven IFN-I production.
- In cutaneous LE (CLE) nonlesional skin, CD16+ DCs — not pDCs — appear to be the active drivers of IFN education, suggesting disease initiation may not be pDC-dependent.
- Non-pDC IFN-I sources (macrophages, keratinocytes, fibroblasts) are clinically significant, and the dominant IFN-producing cell type likely varies by organ, disease stage, and individual patient.
- Three pDC-targeting biologics are in clinical development; functional modulation (litifilimab/BDCA2) may be safer than full depletion (daxdilimab, talacotuzumab) given the role of pDCs in antiviral defense.
- No biologic is yet approved for CLE — an urgent unmet need this therapeutic class is well-positioned to address.
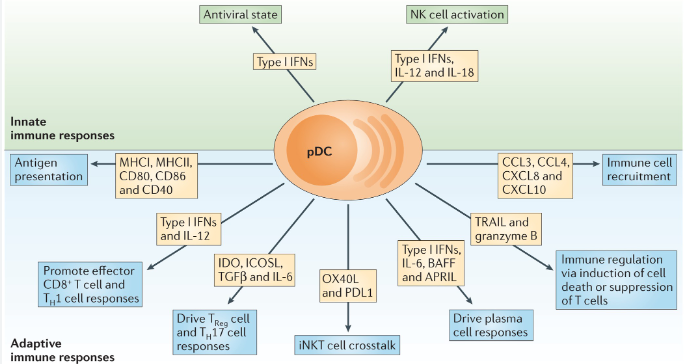
Plasmacytoid dendritic cells (pDCs) have been strongly implicated in the pathogenesis of lupus, as their primary product, type I interferon (IFN-I), drive systemic lupus erythematosus (SLE) and cutaneous lupus erythematosus (CLE). A recent comprehensive review synthesizes the available data on pDCs and, importantly, what we still don't know.
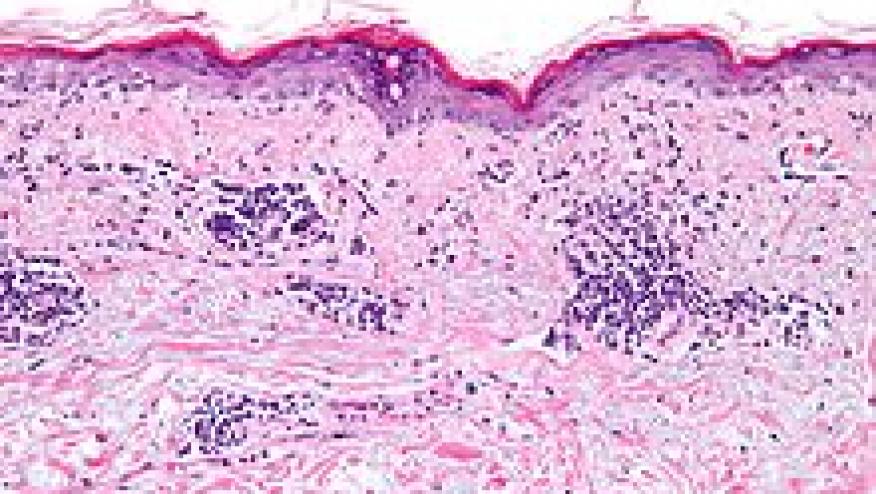
pDCs play a central role in lupus pathogenesis via dysregulated and sustained production of IFN-I, leading to widespread immune activation, chronic inflammation, and organ damage. pDCs migrate (mediated by chemokines, L-selectin, etc) from the blood to peripheral and lesional tissues, where they drive immune responses. pDCs presentation of viral antigens to T cells, under certain conditions, leads to adaptive immune responses. pDC uptake of immune complexes (ICs) can activate TLR7/9 pathways to amplify IFN-I production. Beyond endosomal TLR sensing, activation can also occur through the cGAS-STING pathway. This is important in CLE photosensitivity with UV exposure is cGAS-dependent.
pDCs are checkpoint inhibited or regulated by surface markers like Blood Dendritic Cell Antigen 2 (BDCA2; a C-type lectin receptor expressed exclusively on human pDCs). While pDCs accumulate in lesional skin in both SLE and CLE, the signaling pathways differ: in CLE, tissue inflammation can occur independently of B cells.
This review challenges a pDC-centric dogma in lupus. In nonlesional CLE skin, pDCs are actually predominant but largely inactive, while CD16+ DCs exhibit robust IFN education and active intercellular communication — suggesting CD16+ DCs may play a more important role in disease initiation than previously recognized. Keratinocytes, fibroblasts, macrophages, and neutrophils all contribute meaningfully to the IFN-I signature, and in some CLE biopsies a strong IFN response is present with minimal pDC infiltration.
Targeting pDCs by targeting toll-like receptors (TLRs) is effective in lupus. Hydroxychloroquine, chloroquine, and quinacrine all reduce IFNα production via an inhibitory effect on TLR activation. TYK2 inhibition by deucravacitinib also affects IFN signaling.
There are 3 investigational agents directly targeting pDCs are in development: litifilimab (anti-BDCA2, inhibiting TLR-induced IFN-I production), daxdilimab (anti-ILT7, depleting pDCs), and talacotuzumab (anti-CD123, depleting pDCs).
Litifilimab has shown efficacy in phase 2 trials for both SLE (joints and skin) and CLE, with phase 3 data pending. Daxdilimab achieved near-complete pDC depletion in CLE skin but missed its primary BICLA endpoint in a phase 2 SLE trial. The authors caution that complete pDC depletion risks impairing antiviral immunit. They favor functional modulation over ablation of pDCs.
Nature Reviews Immunology volume 15, pages471–485 (2015)
ADD THE FIRST COMMENT
Disclosures
Disclosures
The author has no conflicts of interest to disclose related to this subject
The author used AI to research and organize this content, and maintains responsibility for its accuracy





If you are a health practitioner, you may Login/Register to comment.
Due to the nature of these comment forums, only health practitioners are allowed to comment at this time.